Презентация по теме «Углеводы и их роль в жизнедеятельности клетки»
Углеводы и их роль в жизнедеятельности клетки
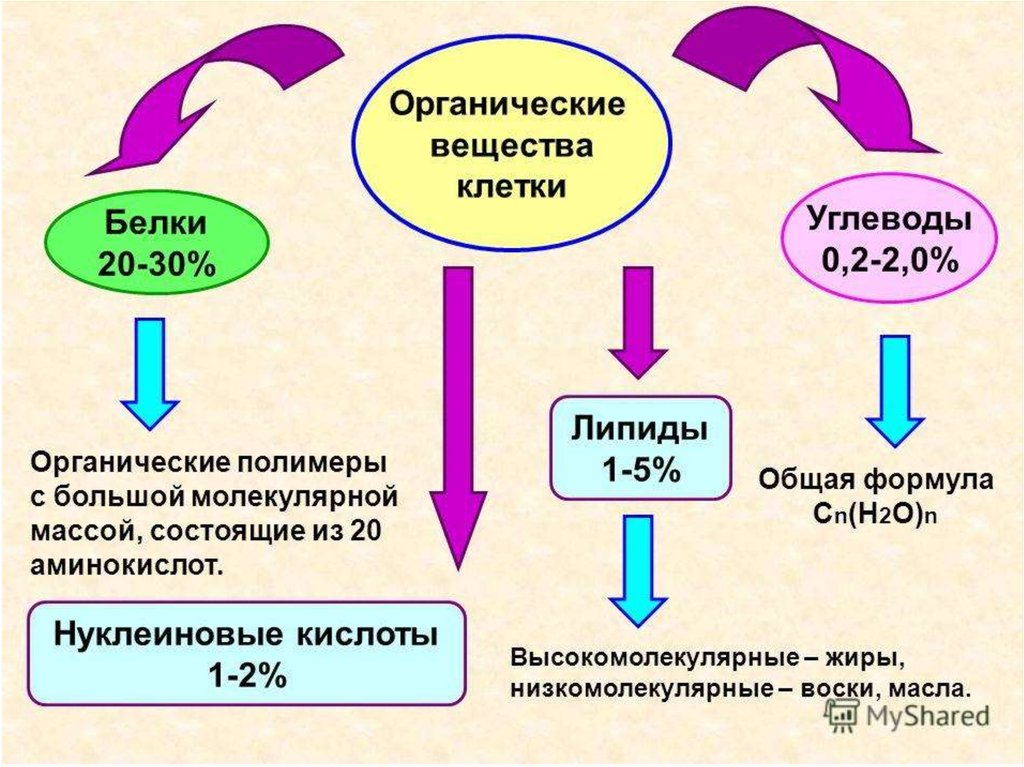
Углеводами называют обширный класс природных органических соединений.
В животных клетках содержится небольшое количество углеводов, а в растительных – почти до 90% от общего количества органических веществ.
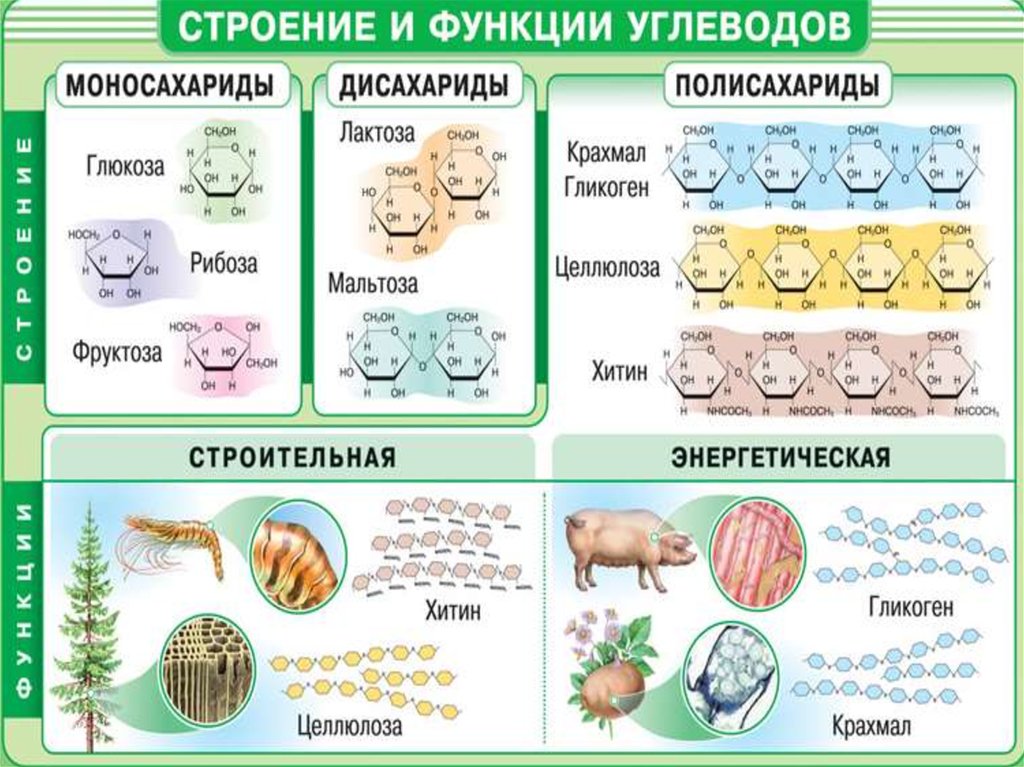
2. КЛАССИФИКАЦИЯ УГЛЕВОДОВ
Олигосахариды
Моносахариды
Это бесцветные вещества, легко растворимые в воде и имеющие сладкий вкус.
К ним относятся:
Тетрозы ( элитроза , треоза)
Пентозы (арабиноза, ксилоза, рибоза)
Гексозы (глюкоза, манноза, галактоза) фруктоза)
Моносахариды
- Рибоза – это простой сахар, натуральное вещество, синтезируемое в организме.
 Рибоза является компонентом нуклеиновых кислот, содержащих генетическую информацию, регулирующих рост, развитие, деление и правильное функционирование клеток.
Рибоза является компонентом нуклеиновых кислот, содержащих генетическую информацию, регулирующих рост, развитие, деление и правильное функционирование клеток. - Дезоксирибоза входит в состав ДНК.
- Галактоза входит в состав некоторых олигосахаридов.
 Рибоза является компонентом нуклеиновых кислот, содержащих генетическую информацию, регулирующих рост, развитие, деление и правильное функционирование клеток.
Рибоза является компонентом нуклеиновых кислот, содержащих генетическую информацию, регулирующих рост, развитие, деление и правильное функционирование клеток.Глюкоза
Глюкозу называют также виноградным сахаром, так как она содержится в большом количестве в виноградном соке. Кроме винограда глюкоза находится и в других сладких плодах и даже в разных частях растений.
Распространена глюкоза и в животном мире: 0,1% ее находится в крови. Глюкоза разносится по всему телу и служит источником энергии для организма. Она также входит в состав сахарозы, лактозы, целлюлозы, крахмала.
Фруктоза
В растительном мире широко распространена фруктоза. Фруктоза содержится в меде. Извлекая из цветов сладких плодов соки, пчелы приготавливают мед, который по химическому составу представляет собой в основном смесь глюкозы и фруктозы.
Фруктоза
Фруктоза содержится в сладких плодах. Также фруктоза входит в состав сложных сахаров, например тростникового и свекловичного.
Д И С А Х А Р И Д Ы —
это сложные сахара, каждая молекула которых при гидролизе распадается на 2 молекулы моносахаридов.
Дисахариды имеют формулу С 12 Н 22 О 11
К дисахаридам относятся:
— с ахароза
— лактоза
— мальтоза
Сахароза
Важнейший из дисахаридов — сахароза — очень распространен в природе. Это химическое название обычного сахара, называемого тростниковым или свекловичным.
Лактоза – молочный сахар
Из молока получают молочный сахар — лактозу. В молоке лактоза содержится в довольно значительном количестве.
В молоке лактоза содержится в довольно значительном количестве.
В молоке млекопитающих лактоза от 4 до 6%.
Лактоза отличается от других сахаров отсутствием гигроскопичности — она не отсыревает.
П О Л И С А Х А Р И Д Ы —
это такие углеводы, которые способны гидролизоваться с образованием множества молекул моносахаридов.
К полисахаридам относятся:
- ( С 6 Н 10 О 5 ) n — целлюлоза (клетчатка),
- (С 6 Н 10 О 5 ) n — крахмал,
— гликоген (животный крахмал),
— хитин.
Полисахариды
- Гликоген содержится во всех животных тканях. Особенно много его в печени (до 20%) и в мышцах (4%).
- Хитин :
* Выполняет защитную и опорную функции, обеспечивая жёсткость клеток — содержится в клеточных стенках грибов.
* Главный компонент экзоскелета членистоногих.
* Также хитин образуется в организмах многих других животных – разнообразных червей, кишечнополостных и т. д.
Крахмал (C 6 Н 10
Целлюлоза является полимером глюкозы.
В ней заключено около 50 % углерода, содержащегося в растениях. По общей массе на Земле целлюлоза занимает первое место среди органических соединений.
- Функции углеводов:
- Углеводы выполняют структурную функцию, то есть участвуют в построении различных клеточных структур (например, клеточных стенок растений) .
- Углеводы выполняют защитную роль у растений (клеточные стенки, состоящие из клеточных стенок мертвых клеток защитные образования — шипы, колючки и др.) .
- Углеводы являются основным энергетическим материалом. При окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.
- Запасающая функция заключается в том, что полисахариды являются запасными питательными веществами всех организмов, играя роль важнейших поставщиков энергии.
Спасибо за внимание!
Углеводы – органические вещества клетки. Биологическая роль углеводов.
Углево́ды (сахариды) —
общее название обширного класса природных
органических соединений. Название
происходит от слов «уголь» и «вода».
Причиной этого является то, что первые
из известных науке углеводов описывались
брутто-формулой Cx(H2O)y,
формально являясь соединениями
Название
происходит от слов «уголь» и «вода».
Причиной этого является то, что первые
из известных науке углеводов описывались
брутто-формулой Cx(H2O)y,
формально являясь соединениями
С точки зрения химии углеводы являются органическими веществами, содержащими неразветвленную цепь из нескольких атомов углерода,карбонильную группу, а также несколько гидроксильных групп.
Биологическое значение углеводов:
Углеводы выполняют структурную функцию, то есть участвуют в построении различных клеточных структур (например, клеточных стенок растений).
Углеводы выполняют защитную роль у растений (клеточные стенки, состоящие из клеточных стенок мертвых клеток защитные образования — шипы, колючки и др.
).Углеводы выполняют
пластическую функцию — хранятся в виде запаса питательных веществ, а также входят в состав сложных молекул (например, пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ, ДНК и РНК.Углеводы являются основным энергетическим материалом. При окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 гводы.
Углеводы участвуют в обеспечении осмотического давления и осморегуляции. Так, в крови содержится 100—110 мг/% глюкозы. От концентрации глюкозы зависит осмотическое давление крови.
Углеводы выполняют рецепторную функцию — многие олигосахариды входят в состав воспринимающей части
 ).
).
В суточном рационе человека и животных преобладают углеводы. Травоядные получают крахмал, клетчатку, сахарозу. Хищники получаютгликоген с мясом.
Уровни
организации живой материи —
иерархически соподчиненные уровни
организации биосистем,
отражающие уровни их усложнения. Чаще
всего выделяют шесть основных структурных
уровней
жизни: молекулярный, клеточный, организменный,популяционно—видовой, биогеоценотический и биосферный.
В типичном случае каждый из этих уровней
является системой из подсистем
нижележащего уровня и подсистемой
системы более высокого уровня.
Методы изучения биологии. Для изучения живой природы биологи применяют различные методы. Наблюдение позволяет выявить объекты и явления.Сравнение дает возможность установить закономерности, общие для разных явлений в живой природе. 8 эксперименте или в опыте создается ситуация, помогающая выявить те или иные свойства биологических объектов. Исторический метод позволяет на основе данных о современном органическом мире и его прошлом познавать процессы развития живой природы. Кроме этих основных методов применяется мно
При
изучении биологических объектов
используется самая различная
техника: микроскопы, ультрацентрифуги,
разнообразные
химические анализаторы, компьютеры и
множество других приборов,
позволяющих раскрыть тайны живой
материи. Свой вклад
в изучение биологии вносят специалисты,
казалось бы, далекие
от биологии: химики, физики, математики,
инженеры и
многие другие.
Свой вклад
в изучение биологии вносят специалисты,
казалось бы, далекие
от биологии: химики, физики, математики,
инженеры и
многие другие.
Различные биологические функции углеводных цепей, полученные из мыши с дефицитом гликозилтрансферазы
1. Allen A.C., Bailey E.M., Barratt J., Buck K.S., Feehally J.1999. Анализ О-гликанов IgA1 в IgA нефропатии с помощью электрофореза углеводов с использованием флуорофора. Дж. Ам. соц. Нефрол. 10: 1763–1771 гг. [PubMed] [Google Scholar]
2. Араи А., Танака К., Икеучи Т., Игараси С., Кобаяши Х., Асака Т., Датэ Х., Сайто М., Танака Х., Кавасаки С. , Уяма Э., Мидзусава Х., Фукухара Н., Цудзи С.2002. Новая мутация в гене GNE и неравновесие по сцеплению в японских родословных. Энн. Нейрол. 52: 516–519. doi: 10.1002/ana.10341 [PubMed] [CrossRef] [Google Scholar]
3. Асано М., Фурукава К., Кидо М., Мацумото С., Умесаки Ю., Кочибе Н., Ивакура Ю. 1997. Задержка роста и ранняя гибель
Мыши с нокаутом бета-1,4-галактозилтрансферазы с усиленной пролиферацией и аномальными
дифференцировка эпителиальных клеток. ЕМБО
Дж. 16: 1850–1857.
doi: 10.1093/emboj/16.8.1850 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ЕМБО
Дж. 16: 1850–1857.
doi: 10.1093/emboj/16.8.1850 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
4. Asano M., Gruss P.1992. Pax-5 выражен в граница между средним и задним мозгом во время развития мыши. Мех. Дев. 39: 29–39. дои: 10.1016/0925-4773(92)-D [PubMed] [CrossRef] [Google Scholar]
5. Асано М., Хаяши М., Йошида Э., Каваде Ю., Ивакура Ю. 1990. Индукция интерферона-альфа интерферона-бета, но не интерферона-бета интерфероном-альфа, в мышь. Вирусология 176: 30–38. дои: 10.1016/0042-6822(90)
-I [PubMed] [CrossRef] [Google Scholar]6. Асано М., Нагата С.1992. Связывание конститутивных и индуцибельных факторов к регуляторному элементу 3 в промоторе гена, кодирующего мышиный гранулоцит колониестимулирующий фактор. Гена 121: 371–375. дои: 10.1016/0378-1119(92)
-G [PubMed] [CrossRef] [Google Scholar]
7. Асано М., Нисидзава М., Нагата С., 1991. Три отдельных регулирующих элемента
промотор положительно активирует транскрипцию мышиного гена, кодирующего
гранулоцитарный колониестимулирующий фактор.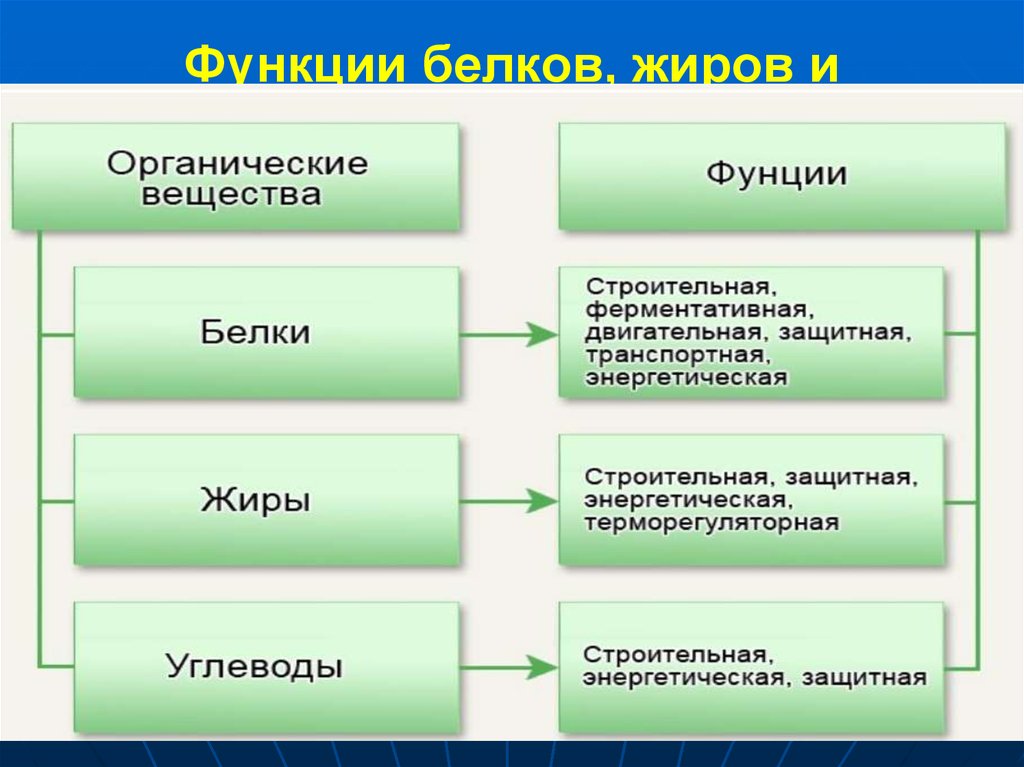 Гена 107:
241–246. дои:
10.1016/0378-1119(91)
Гена 107:
241–246. дои:
10.1016/0378-1119(91)
-5 [PubMed] [CrossRef] [Google Scholar]
8. Чуй Д., О-Эда М., Ляо Ю.Ф., Паннеерсельвам К., Лал А., Марек К.В., Фриз Х.Х., Моремен К.В., Фукуда М.Н., Март Дж.Д., 1997. Результаты дефицита альфа-маннозидазы-II в дизэритропоэзе и раскрывает альтернативный путь образования олигосахаридов биосинтез. Сотовый 90: 157–167. дои: 10.1016/S0092-8674(00)80322-0 [PubMed] [CrossRef] [Google Scholar]
9. Айзенберг И., Авидан Н., Потиха Т., Хохнер Х., Чен М., Олендер Т., Бараш М., Шемеш М., Саде М., Грабов-Нардини Г., Шмилевич И., Фридманн А., Карпати Г., Брэдли В.Г., Баумбах Л., Ланцет Д., Ашер Э.Б., Бекманн Дж.С., Аргов З., Митрани-Розенбаум С.2001. UDP-N-ацетилглюкозамин Ген 2-эпимеразы/N-ацетилманнозаминкиназы мутирован при рецессивном наследственном включении миопатия тела. Нац. Жене. 29: 83–87. doi: 10.1038/ng718 [PubMed] [CrossRef] [Google Scholar]
10. Френетт П.С., Суббарао С., Мазо И.Б.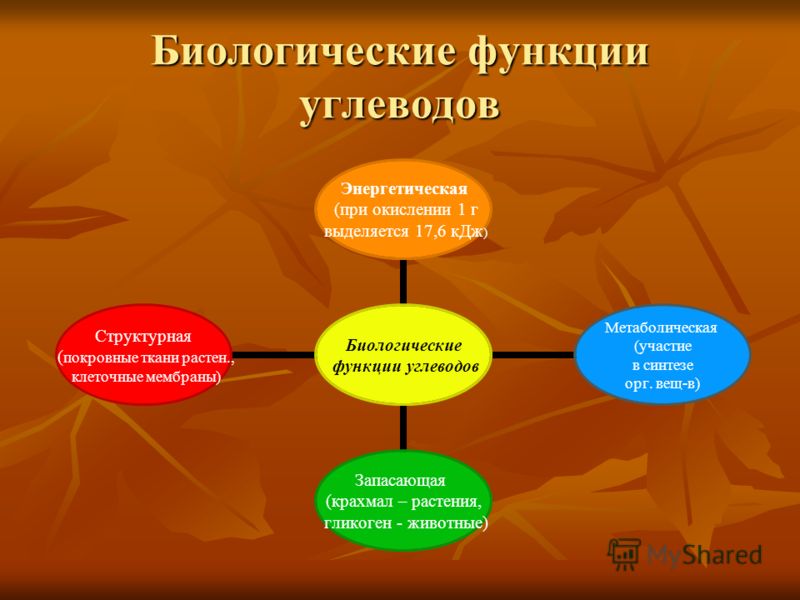 , фон Андриан У.Х., Вагнер Д.Д., 1998. Эндотелиальные селектины и сосудистая клетка
молекула адгезии-1 способствует возвращению гемопоэтических предшественников к кости
костный мозг. Проц. Натл. акад. науч.
США 95: 14423–14428.
doi: 10.1073/pnas.95.24.14423 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, фон Андриан У.Х., Вагнер Д.Д., 1998. Эндотелиальные селектины и сосудистая клетка
молекула адгезии-1 способствует возвращению гемопоэтических предшественников к кости
костный мозг. Проц. Натл. акад. науч.
США 95: 14423–14428.
doi: 10.1073/pnas.95.24.14423 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Granovsky M., Fata J., Pawling J., Muller W.J., Khokha R., Dennis J.W.2000 . Подавление роста опухоли и метастазы у мышей с дефицитом Mgat5. Нац. Мед. 6: 306–312. doi: 10.1038/73163 [PubMed] [CrossRef] [Google Scholar]
12. Hennet T.2002. галактозилтрансфераза семья. Сотовый. Мол. Жизнь науч. 59: 1081–1095. doi: 10.1007/s00018-002-8489-4 [PubMed] [CrossRef] [Google Scholar]
13. Хораи Р., Асано М., Судо К., Канука Х., Судзуки М., Нишихара М., Такахаши М., Ивакура Ю.1998. Получение мышей с дефицитом генов для
интерлейкин (IL)-1альфа, IL-1бета, IL-1альфа/бета и антагонист рецептора IL-1 показывает
что IL-1бета играет решающую роль в развитии скипидарной лихорадки и глюкокортикоидов
секреция. Дж. Эксп.
Мед. 187: 1463–1475.
doi: 10.1084/jem.187.9.1463 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Эксп.
Мед. 187: 1463–1475.
doi: 10.1084/jem.187.9.1463 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Horai R., Saijo S., Tanioka H., Nakae S., Sudo K., Okahara A. ., Икусэ Т., Асано М., Ивакура Ю.2000. Развитие хронического воспалительного артропатия, напоминающая ревматоидный артрит в рецепторе интерлейкина 1 мышей с дефицитом антагонистов. Дж. Экспл. Мед. 191: 313–320. doi: 10.1084/jem.191.2.313 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Иоффе Э., Стэнли П.1994. Мышей не хватает Активность N-ацетилглюкозаминилтрансферазы I снижается в середине беременности, что свидетельствует о существенном роль сложных или гибридных N-связанных углеводов. Проц. Натл. акад. науч. США 91: 728–732. дои: 10.1073/pnas.91.2.728 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Ито М., Сугихара К., Асака Т., Тояма Т., Йошихара Т., Фуруичи К., Вада Т., Асано М.2012. Гипосиалирование гликопротеинов приводит к
к нефротоподобному синдрому, который предотвращается введением сиаловой кислоты при ГНЭ
Мыши с точечной мутацией V572L. ПЛОС
Один 7: e29873. doi: 10.1371/journal.pone.0029873 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ПЛОС
Один 7: e29873. doi: 10.1371/journal.pone.0029873 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Iwakura Y., Asano M., Nishimune Y., Kawade Y.1988. Мужское бесплодие трансгенных мышей несущий экзогенный мышиный ген интерферона-бета под контролем металлотионеина энхансер-промотор. ЕМБО Дж. 7: 3757–3762. doi: 10.1002/j.1460-2075.1988.tb03259.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Kannagi R.2002. Регуляторная роль углеводных лигандов для селектинов в хоуминге лимфоцитов. Курс. мнение Структура биол. 12: 599–608. дои: 10.1016/S0959-440X(02)00365-2 [PubMed] [CrossRef] [Google Scholar]
19. Кумагаи Т., Танака М., Йокояма М., Сато Т., Синкай Т., Фурукава К.2009. Ранняя летальность бета-1,4-галактозилтрансфераза V-мутантных мышей по задержке роста. Биохим. Биофиз. Рез. коммун. 379: 456–459. дои: 10.1016/j.bbrc.2008.12.078 [PubMed] [CrossRef] [Google Scholar]
20. Lowe J. B.2003. Гликанзависимая адгезия лейкоцитов и
рекрутирование при воспалении. Курс. мнение Клетка
биол. 15: 531–538.
doi: 10.1016/j.ceb.2003.08.002 [PubMed] [CrossRef] [Google Scholar]
B.2003. Гликанзависимая адгезия лейкоцитов и
рекрутирование при воспалении. Курс. мнение Клетка
биол. 15: 531–538.
doi: 10.1016/j.ceb.2003.08.002 [PubMed] [CrossRef] [Google Scholar]
21. Lu Q., Hasty P., Shur B.D.1997. Целевая мутация в бета1,4-галактозилтрансфераза приводит к гипофизарной недостаточности и неонатальной летальность. Дев. биол. 181: 257–267. doi: 10.1006/dbio.1996.8444 [PubMed] [CrossRef] [Google Scholar]
22. Marquardt T., Denecke J.2003. Врожденные нарушения гликозилирования: обзор их молекулярных основ, клинических проявлений и конкретных терапии. Евро. Дж. Педиатр. 162: 359–379. doi: 10.1007/s00431-002-1136-0 [PubMed] [CrossRef] [Google Scholar]
23. Мецлер М., Герц А., Саркар М., Шахтер Х., Шредер Дж. В., Март Дж. Д., 1994. Комплекс, связанный с аспарагином
олигосахариды необходимы для морфогенеза во время постимплантационного периода.
разработка. ЕМБО
Дж. 13: 2056–2065.
doi: 10.1002/j.1460-2075.1994. tb06480.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
tb06480.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Mori R., Kondo T., Nishie T., Ohshima T., Asano M. .2004. Нарушение заживления кожных ран в Мыши с дефицитом бета-1,4-галактозилтрансферазы и сниженным уровнем лейкоцитов набор персонала. утра. Дж. Патол. 164: 1303–1314. doi: 10.1016/S0002-9440(10)63217-8 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Nishie T., Hikimochi Y., Zama K., Fukusumi Y., Ito M ., Йокояма Х., Нарусэ К., Ито М., Асано М.2010. Бета4-галактозилтрансфераза-5 представляет собой лактозилцерамидсинтаза, необходимая для экстраэмбрионального развития мышей разработка. Гликобиология 20: 1311–1322 гг. doi: 10.1093/glycob/cwq098 [PubMed] [CrossRef] [Google Scholar]
26. Ниши Т., Мияиси О., Адзума Х., Камеяма А., Нарусэ С., Хасимото Н., Ёкояма Х., Наримацу Х., Вада Т., Асано М.2007. Выработка иммуноглобулина А
нефропатоподобное заболевание при дефиците бета-1,4-галактозилтрансферазы-I
мышей. утра. Дж.
Патол. 170: 447–456.
doi: 10.2353/ajpath.2007.060559 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
утра. Дж.
Патол. 170: 447–456.
doi: 10.2353/ajpath.2007.060559 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Nomura T., Takizawa M., Aoki J., Arai H., Inoue K., Wakisaka E., Йошизука Н., Имокава Г., Домаэ Н., Такио К., Хаттори М., Мацуо Н.1998. Очистка, клонирование кДНК и экспрессия UDP-Gal: глюкозилцерамид бета-1,4-галактозилтрансфераза крысы мозг. Дж. Биол. хим. 273: 13570–13577. doi: 10.1074/jbc.273.22.13570 [PubMed] [CrossRef] [Google Scholar]
28. Пальмитер Р.Д., Бринстер Р.Л., Хаммер Р.Е., Трумбауэр М.Е., Розенфельд М.Г., Бирнберг Н.К., Эванс Р.М.1982. Драматический рост мышей, которые развиваются из яйца, микроинъецированные генами слияния металлотионеина и гормона роста. Природа 300: 611–615. дои: 10.1038/300611a0 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Takagaki S., Yamashita R., Hashimoto N., Sugihara K., Kanari K., Tabata K., Nishie T., Ока С., Мияниши М., Нарусэ К., Асано М. 2019. Галактозильные углеводные остатки на
гемопоэтические стволовые клетки/клетки-предшественники необходимы для самонаведения и приживления к кости
костный мозг. Науч. Респ. 9:
7133. doi: 10.1038/s41598-019-43551-6 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2019. Галактозильные углеводные остатки на
гемопоэтические стволовые клетки/клетки-предшественники необходимы для самонаведения и приживления к кости
костный мозг. Науч. Респ. 9:
7133. doi: 10.1038/s41598-019-43551-6 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Токуда Н., Нумата С., Ли С., Номура Т., Такидзава М. ., Кондо Ю., Ямасита Ю., Хасимото Н., Киёно Т., Урано Т., Фурукава К., Фурукава К.2013. β4GalT6 участвует в синтезе лактозилцерамид с меньшей интенсивностью, чем β4GalT5. Гликобиология 23: 1175–1183 гг. doi: 10.1093/glycob/cwt054 [PubMed] [CrossRef] [Google Scholar]
31. Ямамото С., Ока С., Иноуэ М., Симута М., Манабэ Т., Такахаши Х., Миямото М., Асано М., Сакагами Дж., Судо К., Ивакура Ю., Оно К., Кавасаки Т.2002. Мыши с дефицитом специфических для нервной системы
углеводный эпитоп HNK-1 проявляет нарушение синаптической пластичности и пространственной
обучение. J. Biol.
хим. 277: 27227–27231.
doi: 10.1074/jbc. C200296200 [PubMed] [CrossRef] [Google Scholar]
C200296200 [PubMed] [CrossRef] [Google Scholar]
32. Ёсида А., Кобаяси К., Маня Х., Танигучи К., Кано Х., Мизуно М., Иназу Т., Мицухаши Х., Такахаши С., Такеучи М., Херрманн Р., Штрауб В., Талим Б., Войт Т., Топалоглу Х., Тода Т., Эндо Т.2001. Мышечная дистрофия и миграция нейронов расстройство, вызванное мутациями в гликозилтрансферазе POMGnT1. Дев. Сотовый 1: 717–724. дои: 10.1016/S1534-5807(01)00070-3 [PubMed] [CrossRef] [Google Scholar]
33. Йошихара Т., Сатакэ Х., Нисиэ Т., Окино Н., Хатта Т., Отани Х., Нарусэ С., Судзуки Х., Сугихара К., Камимура Э., Токуда Н., Фурукава К. ., Фуруркава К., Ито М., Асано М.2018. Лактозилцерамидсинтазы, кодируемые Гены B4galt5 и 6 играют ключевую роль в генерации нейронов и образовании миелина у мышей. Генетика PLoS. 14: е1007545. doi: 10.1371/journal.pgen.1007545 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Yoshihara T., Sugihara K., Kizuka Y., Oka S., Asano M.2009. Нарушение обучаемости/памяти и снижение
экспрессия углевода HNK-1 в организме с дефицитом бета4-галактозилтрансферазы-II
мышей. Дж. Биол.
хим. 284: 12550–12561.
doi: 10.1074/jbc.M809188200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Биол.
хим. 284: 12550–12561.
doi: 10.1074/jbc.M809188200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Роль пищевых углеводов в старении организма
Dickinson DJ, Goldstein B (2016) Методы на основе CRISPR для Геномная инженерия Caenorhabditis elegans . Генетика 202 (3): 885–901. doi:10.1534/генетика.115.182162
Артикул КАС пабмед ПабМед Центральный Google ученый
Корси А.К., Вайтман Б., Чалфи М. (2015) Прозрачное окно в биологию: учебник по Caenorhabditis elegans . Генетика 200 (2): 387–407. doi:10.1534/genetics.115.176099
Артикул КАС пабмед ПабМед Центральный Google ученый
Kenyon CJ (2010) Генетика старения.
Природа 464 (7288): 504–512. дои: 10.1038 / природа08980Артикул КАС пабмед Google ученый
Lee Y, An SWA, Artan M, Seo M, Hwang AB, Jeong D-E, Son HG, Hwang W, Lee D, Seo K, Altintas O, Park S, Lee S-JV (2015) Гены и пути которые влияют на продолжительность жизни Caenorhabditis elegans . В: Мори Н., Мук-Юнг И. (ред.) Механизмы старения: продолжительность жизни, метаболизм и старение мозга. Спрингер, Токио, стр. 123–169. дои: 10.1007/978-4-431-55763-0_8
Stiernagle T (2006) Обслуживание C. elegans . Червяк. doi: 10.1895 / wormbook.1.101.1
ПабМед ПабМед Центральный Google ученый
Saltiel AR, Kahn CR (2001)Передача сигналов инсулина и регуляция метаболизма глюкозы и липидов. Природа 414 (6865): 799–806.
дои: 10.1038/414799aАртикул КАС пабмед Google ученый
Altintas O, Park S, Lee SJ (2016)Роль передачи сигналов инсулина/IGF-1 в продолжительности жизни модельных беспозвоночных, C. elegans и D. melanogaster . BMB Rep 49(2):81–92
Статья КАС пабмед ПабМед Центральный Google ученый
Риддл Д.Л., Суонсон М.М., Альберт П.С. (1981) Взаимодействующие гены в формировании личинок нематод дауэра. Природа 290(5808):668–671
Артикул КАС пабмед Google ученый
Morley JF, Morimoto RI (2004) Регуляция продолжительности жизни у Caenorhabditis elegans с помощью фактора теплового шока и молекулярных шаперонов. Mol Biol Cell 15(2):657–664. doi: 10.1091/mbc.E03-07-0532
Артикул КАС пабмед ПабМед Центральный Google ученый
Lee SS, Kennedy S, Tolonen AC, Ruvkun G (2003) Гены-мишени DAF-16, которые контролируют продолжительность жизни и метаболизм C. elegans . Наука 300 (5619): 644–647. дои: 10.1126/наука.1083614
Артикул КАС пабмед Google ученый
Murphy CT, McCarroll SA, Bargmann CI, Fraser A, Kamath RS, Ahringer J, Li H, Kenyon C (2003) Гены, которые действуют после DAF-16, влияя на продолжительность жизни Caenorhabditis elegans . Природа 424 (6946): 277–283. дои: 10.1038 / природа01789
Артикул КАС пабмед Google ученый
Lee SJ, Murphy CT, Kenyon C (2009) Глюкоза сокращает продолжительность жизни C. elegans за счет подавления активности DAF-16/FOXO и экспрессии гена аквапорина. Cell Metab 10 (5): 379–391. doi:10.1016/j.cmet.2009.10.003
Артикул КАС пабмед ПабМед Центральный Google ученый
Ху П.Дж. (2007) Дауэр. Червяк. дои: 10.1895/червьбук.1.144.1
ПабМед ПабМед Центральный Google ученый
Hibuse T, Maeda N, Funahashi T, Yamamoto K, Nagasawa A, Mizunoya W, Kishida K, Inoue K, Kuriyama H, Nakamura T, Fushiki T, Kihara S, Shimomura I (2005) Дефицит аквапорина 7 связан с развитием ожирения за счет активации жировой глицеролкиназы. Proc Natl Acad Sci USA 102(31):10993–10998. doi:10.1073/pnas.0503291102
Артикул КАС пабмед ПабМед Центральный Google ученый
Роек А.М., Сковронски М.Т., Фухтбауэр Э.М., Фухтбауэр А.С., Фентон Р.А., Агре П., Фрокиар Дж., Нильсен С.
(2007) Дефектный метаболизм глицерина у мышей с нокаутом по аквапорину 9 (AQP9). Proc Natl Acad Sci USA 104(9):3609–3614. doi:10.1073/pnas.0610894104Артикул КАС пабмед ПабМед Центральный Google ученый
Pang S, Lynn DA, Lo JY, Paek J, Curran SP (2014) SKN-1 и Nrf2 связывают катаболизм пролина с метаболизмом липидов при недостатке питательных веществ. Община нац. 5:5048. дои: 10.1038/ncomms6048
Артикул КАС пабмед ПабМед Центральный Google ученый
Bishop NA, Guarente L (2007) Два нейрона опосредуют продолжительность жизни, вызванную ограничением диеты, у C. elegans . Природа 447 (7144): 545–549. дои: 10.1038 / природа 05904
Артикул КАС пабмед Google ученый
Walker AK, Näär AM (2012) SREBP: регуляторы холестерина/липидов как терапевтические мишени при нарушениях обмена веществ, раке и вирусных заболеваниях. Clin Lipidol 7(1):27–36
Статья КАС Google ученый
Horton JD, Shimomura I, Brown MS, Hammer RE, Goldstein JL, Shimano H (1998) Активация синтеза холестерина вместо синтеза жирных кислот в печени и жировой ткани трансгенных мышей, гиперпродуцирующих белок, связывающий регуляторный элемент стерола. 2. Дж. Клин Инвест 101 (11): 2331–2339. дои: 10.1172/JCI2961
Артикул КАС пабмед ПабМед Центральный Google ученый
Уокер А.К., Джейкобс Р.Л., Уоттс Дж.Л., Ротьерс В., Цзян К., Финнеган Д.М., Шиода Т., Хансен М., Ян Ф., Нибергалл Л.Дж., Вэнс Д.Е., Цонева М., Харт А.С., Наар А.М. (2011) A законсервировано Цепь обратной связи SREBP-1/фосфатидилхолин регулирует липогенез у многоклеточных животных. Ячейка 147 (4): 840–852. doi:10.1016/j.cell.2011.090,045
Артикул КАС пабмед ПабМед Центральный Google ученый
Уокер А.К., Ян Ф., Цзян К., Джи Дж.И., Уоттс Дж.Л., Пурушотам А., Босс О, Хирш М.Л., Рибич С., Смит Дж.Дж., Исраэлян К., Вестфал К., Роджерс Дж.Т., Шиода Т., Элсон С.Л., Маллиган P, Najafi-Shoushtari H, Black JC, Thakur JK, Kadyk LC, Whetstine JR, Mostoslavsky R, Puigserver P, Li X, Dyson NJ, Hart AC, Naar AM (2010) Сохраняющаяся роль ортологов SIRT1 в зависимом от голодания ингибировании регулятор липидов/холестерина SREBP.
0 Гены Дев 24 (13): 1403–1417. дои: 10.1101/gad.1Артикул КАС пабмед ПабМед Центральный Google ученый
Lee D, Jeong DE, Son HG, Yamaoka Y, Kim H, Seo K, Khan AA, Roh TY, Moon DW, Lee Y, Lee SJ (2015) SREBP и MDT-15 защищают C. elegans от ускоренного старения, вызванного глюкозой, путем предотвращения накопления насыщенных жиров. Гены Дев 29 (23): 2490–2503. дои: 10.1101 / прибл. 266304.115
Артикул КАС пабмед ПабМед Центральный Google ученый
Nomura T, Horikawa M, Shimamura S, Hashimoto T, Sakamoto K (2010) Накопление жира у Caenorhabditis elegans опосредовано гомологом SREBP SBP-1. Гены Нутр 5 (1): 17–27. doi: 10.1007/s12263-009-0157-y
Артикул КАС пабмед Google ученый
Allen BL, Taatjes DJ (2015)Медиаторный комплекс: центральный интегратор транскрипции. Nat Rev Mol Cell Biol 16(3):155–166. дои: 10.1038/nrm3951
Артикул КАС пабмед ПабМед Центральный Google ученый
Taubert S, Van Gilst MR, Hansen M, Yamamoto KR (2006) Медиаторная субъединица, MDT-15, объединяет регуляцию метаболизма жирных кислот с помощью NHR-49.-зависимые и -независимые пути у C.
elegans . Гены Дев 20 (9): 1137–1149. дои: 10.1101/gad.1395406Артикул КАС пабмед ПабМед Центральный Google ученый
Goh GY, Martelli KL, Parhar KS, Kwong AW, Wong MA, Mah A, Hou NS, Taubert S (2014)Консервированная субъединица медиатора MDT-15 необходима для реакции на окислительный стресс у Caenorhabditis elegans . Ячейка старения 13 (1): 70–79. дои: 10.1111/ускорение 12154
Артикул КАС пабмед Google ученый
McGhee JD (2007) Кишечник C. elegans . Червяк. doi: 10.1895 / wormbook.1.133.1
ПабМед ПабМед Центральный Google ученый
Becard D, Hainault I, Azzout-Marniche D, Bertry-Coussot L, Ferre P, Foufelle F (2001) Опосредованная аденовирусом сверхэкспрессия белка-1c, связывающего регуляторный элемент стерола, имитирует действие инсулина на экспрессию генов в печени и гомеостаз глюкозы у мышей с диабетом.
Диабет 50(11):2425–2430Артикул КАС пабмед Google ученый
Takahashi A, Shimano H, Nakagawa Y, Yamamoto T, Motomura K, Matsuzaka T, Sone H, Suzuki H, Toyoshima H, Yamada N (2005) Трансгенные мыши со сверхэкспрессией SREBP-1a под контролем промотора PEPCK проявляют резистентность к инсулину, но не диабет. Биохим Биофиз Акта 1740 (3): 427–433. doi:10.1016/j.bbadis.2004.11.006
Артикул КАС пабмед Google ученый
Poitout V, Robertson RP (2008) Глюколипотоксичность: избыток топлива и дисфункция бета-клеток. Endocr Rev 29 (3): 351–366. дои: 10.1210/er.2007-0023
Артикул КАС пабмед Google ученый
Goodridge AG (1972) Регуляция активности ацетилкоэнзима А-карбоксилазы пальмитоилкоэнзимом А и цитратом.
J Biol Chem 247(21):6946–6952CAS пабмед Google ученый
Ogiwara H, Tanabe T, Nikawa J, Numa S (1978) Ингибирование ацетил-коэнзим-A-карбоксилазы печени крыс пальмитоил-коэнзимом A. Образование эквимолярного комплекса фермент-ингибитор. Eur J Biochem 89(1):33–41
Статья КАС пабмед Google ученый
Раббани Н., Торналли П.Дж. (2015)Дикарбониловый стресс при дисфункции клеток и тканей, способствующий старению и болезням. Biochem Biophys Res Commun 458 (2): 221–226. doi: 10.1016/j.bbrc.2015.01.140
Артикул КАС пабмед Google ученый
Шлоттерер А., Кукудов Г., Бозоргмер Ф., Хаттер Х., Дю Х., Ойконому Д., Ибрагим Ю., Пфистерер Ф., Раббани Н., Торналли П., Сайед А., Флеминг Т., Хамперт П., Швенгер В.
, Зейер М., Хаманн A, Stern D, Brownlee M, Bierhaus A, Nawroth P, Morcos M (2009) C. elegans в качестве модели для изучения сокращения продолжительности жизни, опосредованного высоким уровнем глюкозы. Диабет 58 (11): 2450–2456. дои: 10.2337/db09-0567Артикул КАС пабмед ПабМед Центральный Google ученый
Hardie DG, Ross FA, Hawley SA (2012) AMPK: датчик питательных веществ и энергии, который поддерживает энергетический гомеостаз. Nat Rev Mol Cell Biol 13(4):251–262. дои: 10.1038/nrm3311
Артикул КАС пабмед Google ученый
Mair W, Morantte I, Rodrigues AP, Manning G, Montminy M, Shaw RJ, Dillin A (2011)Увеличение продолжительности жизни, вызванное AMPK и кальциневрином, опосредовано CRTC-1 и CREB. Природа 470 (7334): 404–408. дои: 10.1038 / природа 09706
Артикул КАС пабмед ПабМед Центральный Google ученый
Hwang AB, Ryu EA, Artan M, Chang HW, Kabir MH, Nam HJ, Lee D, Yang JS, Kim S, Mair WB, Lee C, Lee SS, Lee SJ (2014) Регулирование обратной связи через AMPK и HIF-1 опосредует зависимое от АФК долголетие у Caenorhabditis elegans . Proc Natl Acad Sci USA 111(42):E4458–E4467. doi:10.1073/pnas.1411199111
Артикул КАС пабмед ПабМед Центральный Google ученый
Apfeld J, O’Connor G, McDonagh T, DiStefano PS, Curtis R (2004) AMP-активируемая протеинкиназа AAK-2 связывает уровни энергии и инсулиноподобные сигналы с продолжительностью жизни в С.
элегантный . Гены Дев 18 (24): 3004–3009. дои: 10.1101/gad.1255404Артикул КАС пабмед ПабМед Центральный Google ученый
Greer EL, Dowlatshahi D, Banko MR, Villen J, Hoang K, Blanchard D, Gygi SP, Brunet A (2007) Путь AMPK-FOXO опосредует долголетие, индуцированное новым методом диетического ограничения у C. elegans . Curr Biol 17 (19): 1646–1656. doi: 10.1016 / j.cub.2007.08.047
Артикул КАС пабмед ПабМед Центральный Google ученый
Schulz TJ, Zarse K, Voigt A, Urban N, Birringer M, Ristow M (2007) Ограничение потребления глюкозы продлевает Caenorhabditis elegans продолжительность жизни за счет индукции митохондриального дыхания и усиления окислительного стресса. Cell Metab 6 (4): 280–293. doi:10.1016/j.cmet.2007.08.011
Артикул КАС пабмед Google ученый
Choi SS (2011) Диеты с высоким содержанием глюкозы сокращают продолжительность жизни Caenorhabditis elegans за счет индукции эктопического апоптоза. Nutr Res Pract 5 (3): 214–218. doi: 10.4162/nrp.2011.5.3.214
Артикул КАС пабмед ПабМед Центральный Google ученый
Kitaoka S, Morielli AD, Zhao FQ (2013) FGT-1 представляет собой GLUT2-подобный облегчающий переносчик глюкозы у млекопитающих в Caenorhabditis elegans , нарушение работы которого вызывает накопление жира в клетках кишечника. PLoS One 8(6):e68475. doi:10.1371/journal.pone.0068475
Артикул КАС пабмед ПабМед Центральный Google ученый
Свенск Э., Девкота Р., Сталман М., Ранджи П., Раутан М., Магнуссон Ф., Хаммарстен С., Йоханссон М., Борен Дж., Пилон М. (2016) Caenorhabditis elegans PAQR-2 и IGLR-2 защищают от токсичности глюкозы, модулируя липидный состав мембраны. PLoS Genet 12(4):e1005982. doi: 10.1371/journal.pgen.1005982
Артикул пабмед ПабМед Центральный Google ученый
Tauffenberger A, Vaccaro A, Parker JA (2016)Хрупкое увеличение продолжительности жизни за счет диетического митогормезиса у C.
elegans . Старение (Олбани, Нью-Йорк) 8 (1): 50–61. дои: 10.18632/старение.100863Артикул Google ученый
Svensk E, Stahlman M, Andersson CH, Johansson M, Boren J, Pilon M (2013) PAQR-2 регулирует десатурацию жирных кислот во время адаптации к холоду у C. elegans . PLoS Genet 9(9):e1003801. doi:10.1371/journal.pgen.1003801
Артикул КАС пабмед ПабМед Центральный Google ученый
Ивабу М., Окада-Ивабу М., Ямаути Т., Кадоваки Т. (2015) Рецептор адипонектина/адипонектина при заболеваниях и старении. NPJ Aging Mech Dis 1: 15013. doi: 10.1038/npjamd.2015.13
Артикул Google ученый
Дженсен МБ, Джаспер Х (2014)Митохондриальный протеостаз в контроле старения и долголетия.
Cell Metab 20 (2): 214–225. doi:10.1016/j.cmet.2014.05.006Артикул КАС пабмед ПабМед Центральный Google ученый
Tauffenberger A, Vaccaro A, Aulas A, Vande Velde C, Parker JA (2012) Глюкоза задерживает возрастную протеотоксичность. Ячейка старения 11 (5): 856–866. дои: 10.1111/j.1474-9726.2012.00855.х
Артикул КАС пабмед ПабМед Центральный Google ученый
De Virgilio C, Hottiger T, Dominguez J, Boller T, Wiemken A (1994) Роль синтеза трегалозы в приобретении термотолерантности у дрожжей. I. Генетические доказательства того, что трегалоза является термопротектором. Eur J Biochem 219(1–2):179–186
Статья пабмед Google ученый
Ватанабе М., Кикавада Т., Минагава Н.
, Юкухиро Ф., Окуда Т. (2002) Механизм, позволяющий насекомому выживать при полном обезвоживании и экстремальных температурах. J Exp Biol 205 (Pt 18): 2799–2802CAS пабмед Google ученый
Sakurai M, Furuki T, Akao K, Tanaka D, Nakahara Y, Kikawada T, Watanabe M, Okuda T (2008) Витрификация необходима для ангидробиоза у африканского хирономида, Polypedilum vanderplanki . Proc Natl Acad Sci USA 105(13):5093–5098. doi:10.1073/pnas.0706197105
Артикул КАС пабмед ПабМед Центральный Google ученый
Эльбейн А.Д., Пан Ю.Т., Пастушак И., Кэрролл Д. (2003) Новое понимание трегалозы: многофункциональная молекула. Гликобиология 13(4):17R–27R. doi: 10.1093/гликоб/cwg047
Артикул КАС пабмед Google ученый
Mouchiroud L, Molin L, Kasturi P, Triba MN, Dumas ME, Wilson MC, Halestrap AP, Roussel D, Masse I, Dalliere N, Segalat L, Billaud M, Solari F (2011) Дисбаланс пирувата опосредует метаболическое перепрограммирование и имитирует увеличение продолжительности жизни за счет диетического ограничения в Caenorhabditis elegans . Ячейка старения 10 (1): 39–54. doi:10.1111/j.1474-9726.2010.00640.x
Артикул КАС пабмед Google ученый
Cho SC, Park MC, Keam B, Choi JM, Cho Y, Hyun S, Park SC, Lee J (2010) DDS, 4,4′-диаминодифенилсульфон, увеличивает продолжительность жизни организма.
Proc Natl Acad Sci USA 107(45):19326–19331. doi: 10.1073/pnas.1005078107Артикул КАС пабмед ПабМед Центральный Google ученый
Edwards CB, Copes N, Brito AG, Canfield J, Bradshaw PC (2013) Малат и фумарат продлевают жизнь Caenorhabditis elegans . PLoS One 8(3):e58345. doi:10.1371/journal.pone.0058345
Артикул КАС пабмед ПабМед Центральный Google ученый
Чин Р.М., Фу С., Пай М.И., Вернь Л., Хван Х., Дэн Г., Дьеп С., Ломеник Б., Мели В.С., Монсальве Г.К., Ху Э., Уилан С.А., Ван Д.С., Юнг Г., Солис Г.М., Фазлоллахи F, Kaweeteerawat C, Quach A, Nili M, Krall AS, Godwin HA, Chang HR, Faull KF, Guo F, Jiang M, Trauger SA, Saghatelian A, Braas D, Christofk HR, Clarke CF, Teitell MA, Petrascheck M, Reue K, Jung ME, Frand AR, Huang J (2014)Метаболит альфа-кетоглутарат продлевает продолжительность жизни за счет ингибирования АТФ-синтазы и TOR.
Природа 510(7505):397–401. дои: 10.1038 / природа13264КАС пабмед ПабМед Центральный Google ученый
Denzel MS, Storm NJ, Gutschmidt A, Baddi R, Hinze Y, Jarosch E, Sommer T, Hoppe T, Antebi A (2014) Метаболиты гексозаминового пути улучшают контроль качества белка и продлевают жизнь. Ячейка 156 (6): 1167–1178. doi:10.1016/j.cell.2014.01.061
Артикул КАС пабмед Google ученый
Roux AE, Leroux A, Alaamery MA, Hoffman CS, Chartrand P, Ferbeyre G, Rokeach LA (2009)Прогрессирующие эффекты передачи сигналов глюкозы через G-белок, связанный с глюкозным рецептором у делящихся дрожжей. Генетика PLoS 5(3):e1000408. doi:10.1371/journal.pgen.1000408
Артикул пабмед ПабМед Центральный Google ученый
Семчишин Х.М., Лозинская Л.М., Медзобродский Ю., Лущак В.И. (2011) Фруктоза и глюкоза по-разному влияют на параметры старения и карбонильного/окислительного стресса в клетках Saccharomyces cerevisiae . Carbohydr Res 346 (7): 933–938. doi:10.1016/j.carres.2011.03.005
Артикул КАС пабмед Google ученый
Suarez G, Rajaram R, Oronsky AL, Gawinowicz MA (1989) Неферментативное гликирование бычьего сывороточного альбумина фруктозой (фруктация).
Сравнение с реакцией Майяра, инициируемой глюкозой. J Biol Chem 264(7):3674–3679CAS пабмед Google ученый
Mortuza R, Chen S, Feng B, Sen S, Chakrabarti S (2013)Вызванное высоким уровнем глюкозы изменение SIRT в эндотелиальных клетках вызывает быстрое старение в регулируемом пути p300 и FOXO. PLoS One 8(1):e54514. doi:10.1371/journal.pone.0054514
Артикул КАС пабмед ПабМед Центральный Google ученый
Zhang B, Cui S, Bai X, Zhuo L, Sun X, Hong Q, Fu B, Wang J, Chen X, Cai G (2013) Сверхэкспрессия SIRT3 противодействует ускоренному высоким содержанием глюкозы клеточному старению в диплоидных фибробластах человека через Сигнальный путь SIRT3-FOXO1. Эпоха (Дордр) 35(6):2237–2253. doi: 10.1007/s11357-013-9520-4
Артикул КАС Google ученый
Kuki S, Imanishi T, Kobayashi K, Matsuo Y, Obana M, Akasaka T (2006)Гипергликемия ускорила старение эндотелиальных клеток-предшественников за счет активации митоген-активируемой протеинкиназы p38. Циркуляр J 70 (8): 1076–1081
Артикул КАС пабмед Google ученый
Samaha FF, Iqbal N, Seshadri P, Chicano KL, Daily DA, McGrory J, Williams T, Williams M, Gracely EJ, Stern L (2003) Низкоуглеводная диета по сравнению с диетой с низким содержанием жиров при тяжелой ожирение. N Engl J Med 348 (21): 2074–2081. дои: 10.1056/NEJMoa022637
Артикул КАС пабмед Google ученый
 Природа 464 (7288): 504–512. дои: 10.1038 / природа08980
Природа 464 (7288): 504–512. дои: 10.1038 / природа08980 дои: 10.1038/414799a
дои: 10.1038/414799a «>
«>Hsu AL, Murphy CT, Kenyon C (2003) Регуляция старения и возрастных заболеваний с помощью DAF-16 и фактора теплового шока. Наука 300 (5622): 1142–1145. дои: 10.1126/наука.1083701
Артикул КАС пабмед Google ученый
 «>
«>Tullet JM, Hertweck M, An JH, Baker J, Hwang JY, Liu S, Oliveira RP, Baumeister R, Blackwell TK (2008) Прямое ингибирование фактора долголетия SKN-1 посредством инсулиноподобной передачи сигналов в С. Элеганс . Ячейка 132 (6): 1025–1038. doi:10.1016/j.cell.2008.01.030
Артикул КАС пабмед ПабМед Центральный Google ученый
 «>
«>Murphy CT, Lee SJ, Kenyon C (2007) Увлечение тканей путем обратной регуляции экспрессии гена инсулина в энтодерме Caenorhabditis elegans . Proc Natl Acad Sci USA 104 (48): 19046–19050. doi:10.1073/pnas.0709613104
Артикул КАС пабмед ПабМед Центральный Google ученый
 (2007) Дефектный метаболизм глицерина у мышей с нокаутом по аквапорину 9 (AQP9). Proc Natl Acad Sci USA 104(9):3609–3614. doi:10.1073/pnas.0610894104
(2007) Дефектный метаболизм глицерина у мышей с нокаутом по аквапорину 9 (AQP9). Proc Natl Acad Sci USA 104(9):3609–3614. doi:10.1073/pnas.0610894104 «>
«>Фонтана Л., Партридж Л. (2015) Укрепление здоровья и долголетия с помощью диеты: от модельных организмов до человека. Ячейка 161 (1): 106–118. doi:10.1016/j.cell.2015.02.020
Артикул КАС пабмед ПабМед Центральный Google ученый
 «>
«>McKay RM, McKay JP, Avery L, Graff JM (2003) C elegans : модель для изучения генетики накопления жира. Dev Cell 4(1):131–142
Статья КАС пабмед ПабМед Центральный Google ученый
 Гены Дев 24 (13): 1403–1417. дои: 10.1101/gad.1
Гены Дев 24 (13): 1403–1417. дои: 10.1101/gad.1 «>
«>Ян Ф., Воут Б.В., Саттерли Дж.С., Уокер А.К., Джим Сун З.И., Уоттс Дж.Л., ДеБомон Р., Сайто Р.М., Хайбертс С.Г., Ян С., Макол С., Айер Л., Тиан Р., ван ден Хьювел С., Харт AC, Wagner G, Naar AM (2006)Субъединица ARC / медиатора, необходимая для SREBP-контроля холестерина и гомеостаза липидов. Природа 442 (7103): 700–704. дои: 10.1038 / природа04942
Артикул КАС пабмед Google ученый
 elegans . Гены Дев 20 (9): 1137–1149. дои: 10.1101/gad.1395406
elegans . Гены Дев 20 (9): 1137–1149. дои: 10.1101/gad.1395406 Диабет 50(11):2425–2430
Диабет 50(11):2425–2430 J Biol Chem 247(21):6946–6952
J Biol Chem 247(21):6946–6952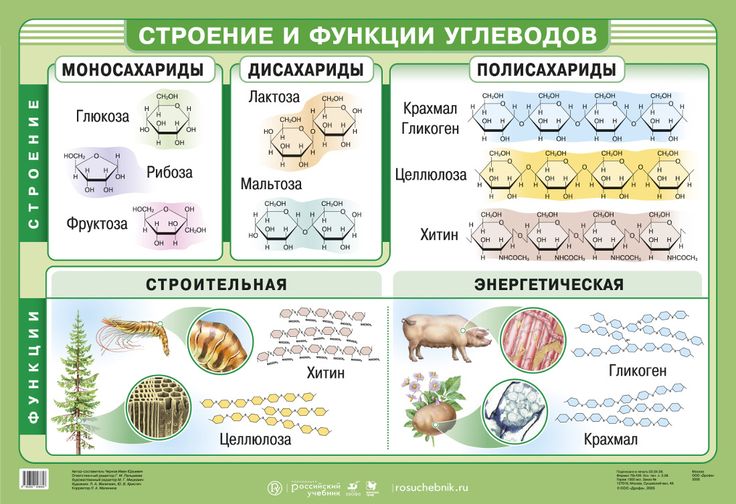 , Зейер М., Хаманн A, Stern D, Brownlee M, Bierhaus A, Nawroth P, Morcos M (2009) C. elegans в качестве модели для изучения сокращения продолжительности жизни, опосредованного высоким уровнем глюкозы. Диабет 58 (11): 2450–2456. дои: 10.2337/db09-0567
, Зейер М., Хаманн A, Stern D, Brownlee M, Bierhaus A, Nawroth P, Morcos M (2009) C. elegans в качестве модели для изучения сокращения продолжительности жизни, опосредованного высоким уровнем глюкозы. Диабет 58 (11): 2450–2456. дои: 10.2337/db09-0567 «>
«>Саги Д., Ким С.К. (2012) Инженерный подход к увеличению срока службы C. elegans . Генетика PLoS 8(6):e1002780. doi:10.1371/journal.pgen.1002780
Артикул КАС пабмед ПабМед Центральный Google ученый
 элегантный . Гены Дев 18 (24): 3004–3009. дои: 10.1101/gad.1255404
элегантный . Гены Дев 18 (24): 3004–3009. дои: 10.1101/gad.1255404 «>
«>Юн Дж., Финкель Т. (2014) Митогормезис. Cell Metab 19 (5): 757–766. doi:10.1016/j.cmet.2014.01.011
Артикул КАС пабмед ПабМед Центральный Google ученый
 «>
«>Feng Y, Williams BG, Koumanov F, Wolstenholme AJ, Holman GD (2013) FGT-1 является основным переносчиком глюкозы у C. elegans и занимает центральное место в путях старения. Биохим J 456 (2): 219–229. дои: 10.1042/BJ20131101
Артикул КАС пабмед ПабМед Центральный Google ученый
 elegans . Старение (Олбани, Нью-Йорк) 8 (1): 50–61. дои: 10.18632/старение.100863
elegans . Старение (Олбани, Нью-Йорк) 8 (1): 50–61. дои: 10.18632/старение.100863 Cell Metab 20 (2): 214–225. doi:10.1016/j.cmet.2014.05.006
Cell Metab 20 (2): 214–225. doi:10.1016/j.cmet.2014.05.006 , Юкухиро Ф., Окуда Т. (2002) Механизм, позволяющий насекомому выживать при полном обезвоживании и экстремальных температурах. J Exp Biol 205 (Pt 18): 2799–2802
, Юкухиро Ф., Окуда Т. (2002) Механизм, позволяющий насекомому выживать при полном обезвоживании и экстремальных температурах. J Exp Biol 205 (Pt 18): 2799–2802 «>
«>Honda Y, Tanaka M, Honda S (2010) Трегалоза продлевает жизнь нематоде Caenorhabditis elegans . Ячейка старения 9 (4): 558–569. doi:10.1111/j.1474-9726.2010.00582.x
Артикул КАС пабмед Google ученый
 Proc Natl Acad Sci USA 107(45):19326–19331. doi: 10.1073/pnas.1005078107
Proc Natl Acad Sci USA 107(45):19326–19331. doi: 10.1073/pnas.1005078107 Природа 510(7505):397–401. дои: 10.1038 / природа13264
Природа 510(7505):397–401. дои: 10.1038 / природа13264 «>
«>Вайнбергер М., Мескита А., Кэролл Т., Маркс Л., Ян Х., Чжан З., Людовико П., Бурханс В.К. (2010) Передача сигналов роста способствует хронологическому старению у почкующихся дрожжей за счет индукции анионов супероксида, которые ингибируют покой. Старение (Олбани, Нью-Йорк) 2 (10): 709–726. дои: 10.18632/старение.100215
Артикул КАС Google ученый
 Сравнение с реакцией Майяра, инициируемой глюкозой. J Biol Chem 264(7):3674–3679
Сравнение с реакцией Майяра, инициируемой глюкозой. J Biol Chem 264(7):3674–3679 «>
«>Imai S, Guarente L (2014) NAD+ и сиртуины при старении и болезнях. Тенденции Cell Biol 24 (8): 464–471. doi: 10.1016/j.tcb.2014.04.002
Артикул КАС пабмед ПабМед Центральный Google ученый
